|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Le dispositif distributeur de pollen des étamines matures
de Salvia pratensis
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
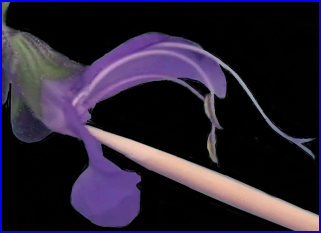
Le suivi du développement des étamines de la sauge permet de comprendre les particularités de leur structure. A maturité, elles forment un dispositif singulier capable de délivrer du pollen à un transporteur (bourdon ou une abeille) quand le butineur récolte le nectar.
Comment le dispositif staminal fonctionne-t-il?
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Les observations sur le terrain (voir vidéo introductive) montrent que le bourdon récoltant le nectar au fond de la corolle de Salvia pratensis déclenche l'abaissement des étamines qui déposent du pollen sur son dos.
On peut facilement simuler l'intervention de l'insecte et comprendre comment le mouvement des étamines est initié (vidéo ci-dessous). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
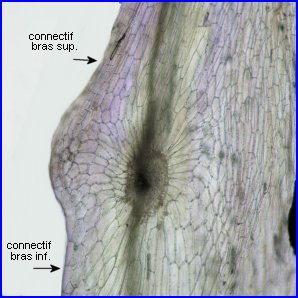
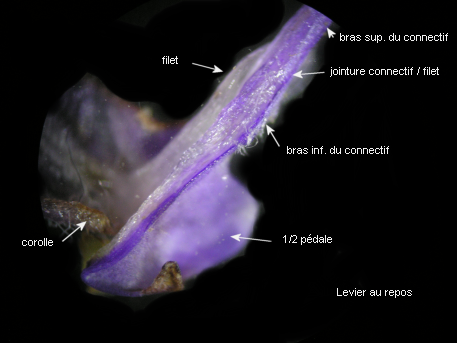
Si on exerce une poussée suffisante sur les parties inférieures élargies et solidarisées des connectifs staminaux, les bras supérieurs basculent vers l'avant entraînant dans leur mouvement les loges polliniques. Dès que la pression cesse, les étamines reprennent leur position initiale.
La partie sur laquelle est exercée la poussée fonctionne comme une pédale. Le pivotement a lieu autour des points de contact entre les filets et les connectifs qui ont donc un rôle central dans le fonctionnement du levier. Les courts connectifs, sont solidement fixés à la corolle. |
|
|
|
|
|
|
|
|
|
|
|
|
Morphologie du dispositif staminal (mature) distributeur de pollen |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
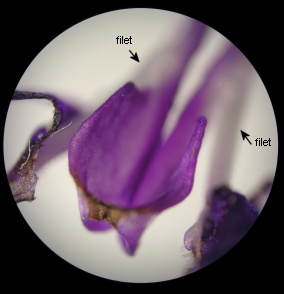
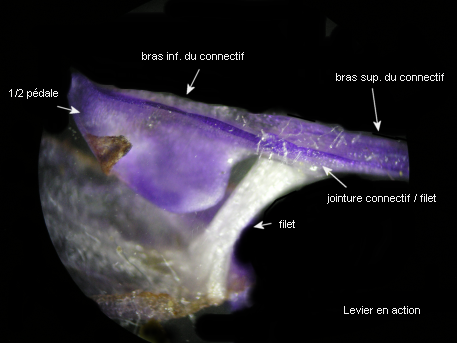
La pédale du levier est située juste à l'entrée du tube de la corolle, entre les filets, et l'obstrue en grande partie. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Dans la fleur mature (image à gauche), au stade mâle, la loge polllinique fertile unique, déhiscente, de chaque étamine est cachée dans le casque de la lèvre supérieure de la corolle. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
La pédale |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
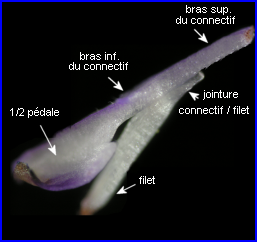
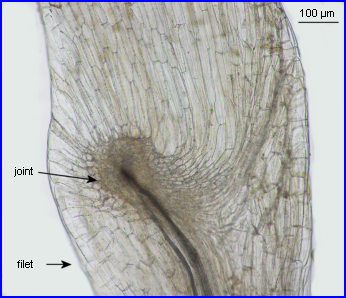
Les courts filets des étamines ancrent le dispositif staminal à la corolle par leur extrémité inférieure, l'autre formant une jonction élastique avec le connectif.
L'image montre bien comment les parties élargies du connectif, porteuses chacune, à l'origine d'une loge pollinique avortée, sont solidarisées et forment une pédale mobile (balancier) avec un dispositif en anneau ménageant un étroit passage. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
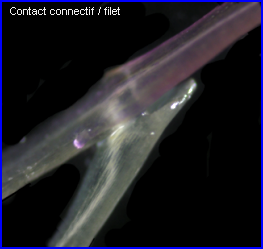
| Le joint filet / connectif |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
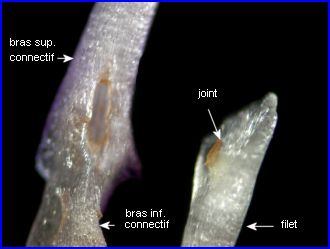
Pour comprendre l'organisation de la jointure entre le connectif et le filet, il est indispensable d'observer les 2 pièces staminales après les avoir écartées puis séparées. |
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
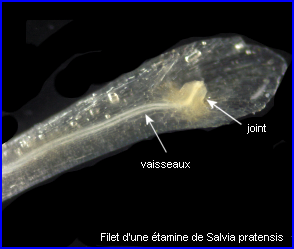
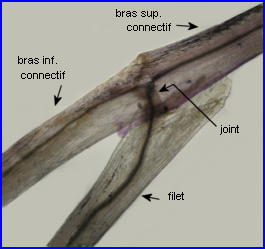
Connectif et filet sont réunis par un joint très court parcouru par des tissus conducteurs. C'est ce joint qui est actif lors du mouvement de l'étamine.
Une étude histologique du joint est possible. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
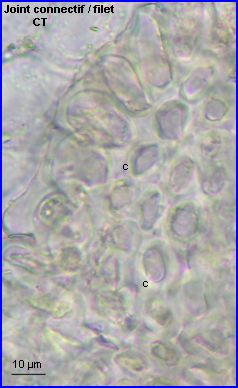
Du côté connectif, on repère la zone de contact du joint avec au centre les tissus conducteurs et à la périphérie un épiderme à grandes cellules. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Le joint est resté accroché au filet, il est limité par un épiderme. Les tissus conducteurs du filet empruntent la partie centrale du joint. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
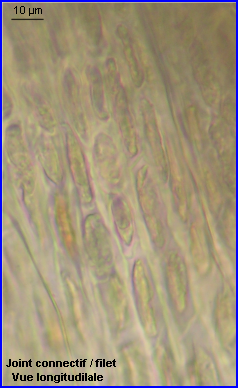
Sous l'épiderme du joint, on trouve un tissu de type collenchyme mais avec des espaces entre certaines cellules. Sur l'image de gauche on observe les sections des cellules du collenchyme (c) alors que sur celle de droite, ce sont les files de cellules vues longitudinalement qui apparaissent.
Le collenchyme est un tissu de soutien vivant, la paroi épaisse restée cellulosique lui confère à la fois une bonne résistance aux flexions et torsions et une certaine élasticité permettant au joint de conserver une bonne souplesse. |
|
|
|
|
Approche biomécanique du fonctionnement du dispositif staminal distributeur de pollen |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Ces images permettent de bien situer le joint qui relie le connectif et le filet d'une étamine de Salvia pratensis et, à la lumière de la description qui en est faite ci-dessus, on peut essayer d'expliquer comment il intervient dans le mouvement des étamines. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Au repos, les bras inférieurs des connectifs des étamines ainsi que leurs extrémités élargies et solidarisées (pédale) sont situés entre les filets qui fixent l'ensemble du système à la corolle. Les étamines sont alors relevées. Si une poussée suffisante est exercée sur la pédale, une partie de l'énergie est transférée aux joints qui relient chaque connectif au filet correspondant ce qui provoque leur torsion (voir histologie). La base étant bien fixée à la corolle, la torsion des joints entraîne un mouvement de basculement des bras inférieurs stériles et de la pédale vers l'arrière, alors que les bras supérieurs, fertiles basculent vers l'avant avec leur loge pollinique.
Si la pression cesse, l'élasticité des joints (voir histologie) leur permet de reprendre leur forme et leur position initiales, les étamines reprennent alors leur position . Le levier peut fonctionner ainsi plusieurs fois de suite. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|