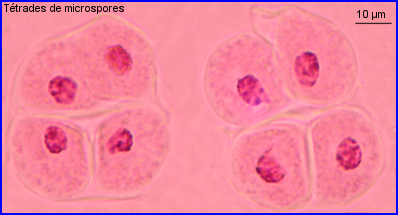
Formation du pollen de l'Arum maculatum
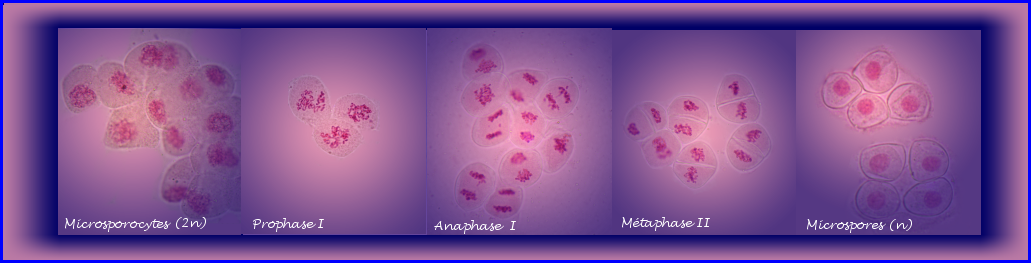
La microsporogenèse
Les fleurs mâles portées par le spadice sont réduites aux anthères des étamines. C'est dans les anthères en cours de développement que les cellules mères du pollen* diploïdes subissent la méiose. Elle conduit aux microspores haploïdes.
Pour observer des figures de méiose, il faut récupérer de jeunes spadices en les débarrassant des feuilles qui les enveloppent. Il faut ensuite réaliser des préparations avec des étamines détachées de l'axe floral.
Ici, les préparations sont colorées à l'orcéine acétique, sans fixation préalable. Elles ne peuvent pas servir à découvrir les phénomènes chromosomiques de la méiose comme on peut le faire, par exemple, avec les préparations de testicules de criquet (voir tp), car le nombre de chromosomes est trop élevé (2n = 56).
L'étude peut cependant permettre de réinvestir des connaissances acquises avec d'autres supports pour reconstituer, à partir des observations réalisées, la chronologie des évènements qui caractérisent le passage de la cellule mère aux 4 microspores.
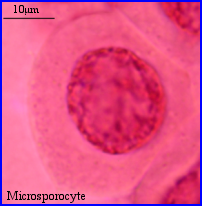
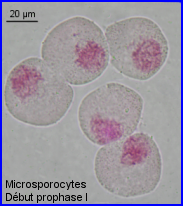
Dans l'anthère immature, les cellules mères du pollen (ou mieux cellules mères des microspores) = microsporocytes ou encore méiocytes, forment quatre massifs, un par sac pollinique. Pour étudier la méiose, on peut séparer les cellules intéressantes du reste de l'anthère en écrasant légèrement (squash) la préparation.
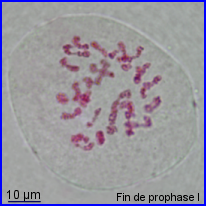
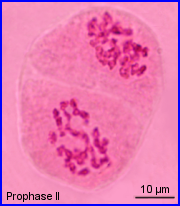
- Prophase I
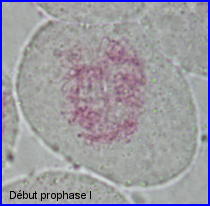
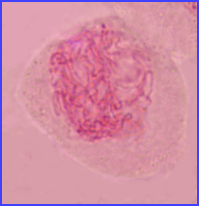
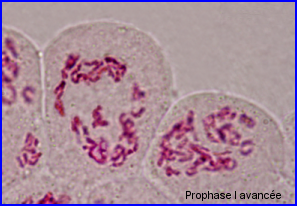
Ils sont fins et longs
Appariement en cours
Les chromosomes homologues sont appariés (bivalents).
28 bivalents juste avant leur migration en plaque équatoriale
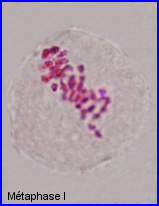
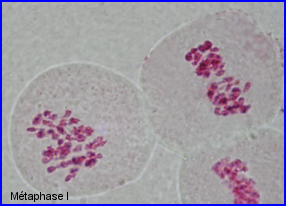
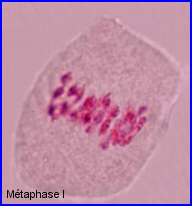
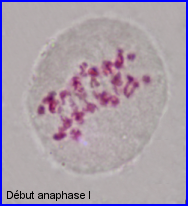
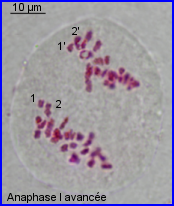
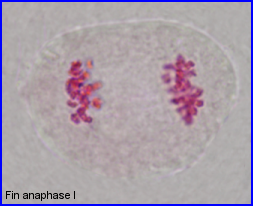
Séparation des chromosomes homologues encore très condensés. Au sein de chaque paire, l'un migre vers un pôle et l'autre vers le pôle opposé.
Séparation de 2 lots haploïdes de n =28 chr. à 2 chromatides chacun. 1 et 1' = chr. homologues
2 et 2' = autres chr. homologues
Ascension polaire terminée.
Début du tassement des chromosomes aux pôles
regroupés dans chaque lot haploïde
Cytocinèse en cours
Edification de la nouvelle cloison.
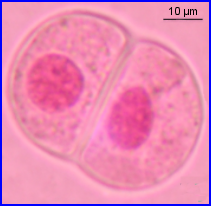
Noyaux reconstitués
2 cellules filles à n chr.à 2 chromatides chacun, décondensés (état chromatine)
Après une courte intercinèse (sans réplication d'ADN), chaque cellule est le siège d'une division bien plus rapide que la précédente.
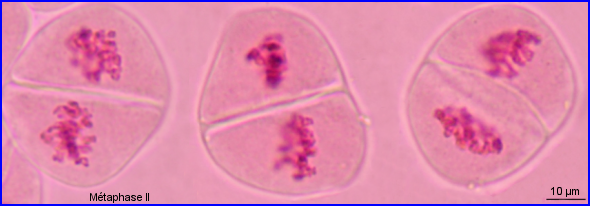
Les préparations n'ont permis d'observer ni l'anaphase II ni la télophase II.
A l'anaphase II, chaque chromosome se partage en 2 chromosomes fils à une chromatide, chacun migrant vers un pôle opposé de la cellule. Il se forme donc 2 lots de n= 28 chr. par cellule. A la télophase II, une enveloppe nucléaire se forme autour de chaque lot haploïde de chromosomes qui se décondensent et retournent à l'état de chromatine et il y a cytocinèse.
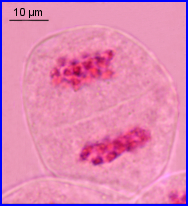
Résultat de la méiose
A partir d'une cellule mère diploïde (microsporocyte), la méiose conduit à 4 microspores haploïdes qui restent groupées et forment une tétrade.