La respiration alternative résistante au cyanure
- Les expérimentations réalisées avec des suspensions de mitochondries végétales dont on suit par oxymétrie la consommation d'O2 (voir résultats) montrent l'existence d'une respiration "classique" inhibée par le cyanure et d'une respiration alternative, résistante au cyanure et inhibée par SHAM. Dans la mitochondrie des plantes, les électrons fournis par les donneurs NADH et succinate peuvent être transférés à O2 (accepteur) soit par la chaîne des cytochromes dont le dernier transporteur est la cytochrome C oxydase (inhibée par les ions CN-
) soit par une chaîne dérivée dont le dernier transporteur est l'AOX (oxydase alternative) insensible aux ions CN- mais inhibée par SHAM.
Rq/ L'oxydation du succinate en fumarate est une réaction du cycle de Krebs catalysée par la succinate déshydrogénase, oxydoréductase qui fait également partie de la chaîne respiratoire. C'est le FAD, coenzyme fixé (liaison covalente) à la succinate déshydrogénase qui accepte les électrons et protons cédés par le succinate et prend la forme réduite FADH2 .
- Les expérimentations montrent également (voir résultats) que dans les mitochondries des organes végétaux non thermogéniques, la respiration alternative est le plus souvent peu intense, voire inexistante alors que dans celle de l'appendice de l'Arum en phase thermogénique elle assure pratiquement seule la consommation d'O2 qui est d'ailleurs très élevée. C'est donc la respiration alternative qui est responsable de l'échauffement de l'appendice lors de la phase thermogénique. Pour comprendre comment elle peut générer autant de chaleur, il faut replacer l'intervention de l'AOX dans le fonctionnement global de la mitochondrie et s'intéresser au couplage transferts d'électrons / phosphorylations oxydatives.
La respiration alternative : un processus biochimique fortement thermogénique
- Le couplage entre oxydoréductions de la chaîne respiratoire "classique" et phosphorylations oxydatives
Les coenzymes NADH, H+ et FADH2 (RH2) issus du catabolisme cellulaire ont un pouvoir réducteur élevé. Chaque molécule peut facilement céder 2 électrons (+ 2 protons) à fort potentiel énergétique et reprendre sa forme oxydée. Dans la mitochondrie leur oxydation commence par séparer les électrons (e-) et protons qu'ils cèdent. Les électrons sont pris en charge par une chaîne de transporteurs (situés dans la membrane interne de la mitochondrie) et transférés par étapes successives (plusieurs oxydoréductions) jusqu'à la cytochrome C oxydase qui les cède à son tour à l'accepteur final O2 alors réduit en H2O. Les électrons sont alors au niveau énergétique le plus bas.
L'énergie chimique initialement présente dans les molécules RH2 est libérée par étapes (et non en une seule fois), chaque oxydoréduction en libérant une quantité plus ou moins importante. L'énergie libérée par certaines oxydoréductions (exergoniques) peut être utilisée pour la synthèse d'ATP (par phosphorylation de l'ADP). Cette réaction énergétiquement défavorable (endergonique) nécessite un apport d'énergie que 3 sites d'oxydoréduction de la chaîne respiratoire peuvent fournir dans le cas de l'oxydation du NADH, H+(seulement 2 dans le cas de celle de FADH2). Ainsi une bonne partie de l'énergie de l'oxydation des petites molécules NADH, H+ et FADH2 est convertie en une énergie chimique stockée temporairement dans la liaison phosphate de l'ATP, le reste est dissipé sous forme de chaleur.
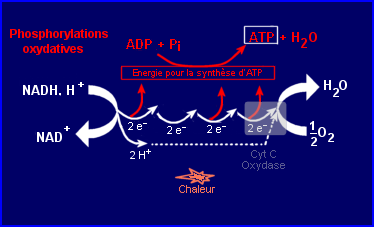
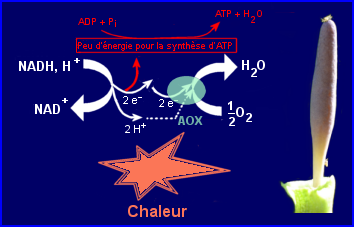
Couplage avec importante synthèse d'ATP
Peu de chaleur dissipée
Peu ou pas de couplage avec synthèse d'ATP
Chaleur dissipée importante
- La respiration alternative contribue peu ou pas à la synthèse d'ATP
La respiration alternative nécessite l'intervention d'une oxydase finale différente de la cytochrome C oxydase, il s'agit de l'alternative oxydase (AOX) enzyme ubiquitaire chez les plantes. Elle est intégrée à la membrane interne de la mitochondrie mais du côté matrice. Elle intervient dans la chaîne respiratoire en recevant des électrons de l'un des transporteurs intermédiaires pour les transférer à O2 qu'elle réduit en H2O. Sa mise en oeuvre court-circuite 2 sites de la chaîne respiratoire qui participent normalement à la conversion de l'énergie d'oxydation du NADH, H+ (ou de FADH2) en énergie utilisable pour la synthèse d'ATP. L'AOX n'est pas inhibée par le cyanure.
Rq/ L'oxydation de FADH2 par la voie de l'AOX n'entraîne pas du tout de synthèse d'ATP. Il existe dans la membrane interne des mitochondries des plantes (et non chez les animaux) une NADH déshydrogénase externe du côté de l'espace intermembranaire et une autre dite interne du côté matrice qui sont différentes de celle qui intervient dans la chaîne respiratoire habituelle. L'intervention de l'une ou l'autre complétée par celle de l'AOX permet une oxydation du NADH, H+ complètement découplée de la synthèse d'ATP.
Au cours de la respiration alternative l'énergie de l'oxydation du NADH n'est que peu ou pas du tout convertie en énergie de liaison phosphate retrouvée dans l'ATP, elle est donc en grande partie ou complètement dissipée sous forme de chaleur. L'énergie de l'oxydation de FADH2 est entièrement dissipée sous forme de chaleur.
Mécanisme de la thermogenèse dans l'appendice de l'Arum maculatum
- Dans les mitochondries végétales, les 2 voies de transport d'électrons peuvent fonctionner simultanément. En général l'AOX n'est pas (ou est peu) exprimée mais dans certaines conditions physiologiques particulières (surcharge énergétique, stress abiotiques ou biotiques) son expression augmente, la voie de l'AOX pouvant alors participer activement à l'oxydation des coenzymes NADH, H+ et FADH2 sans pour cela entraîner un échauffement significatif de l'organe concerné (intensité respiratoire trop faible).
-
Durant le développement de l'appendice de l'Arum maculatum, le nombre de mitochondries par cellule augmente, elles deviennent plus volumineuses et leurs crêtes occupent un volume plus important (H. Mohr and al.). Au moment de la crise respiratoire, la vitesse de la glycolyse (alimentée par la dégradation des réserves d'amidon) qui se déroule dans le cytosol des cellules est très élevée, dans les mitochondries les enzymes du cycle de Krebs sont abondantes, l'AOX est présente en quantité importante, la cytochrome oxydase est détectable (H. Mohr and al.). Le catabolisme glucidique (glycolyse , cycle de Kebs) fournit des transporteurs d'électrons réduits NADH, H+ et FADH2 en grande quantité aux mitochondries nombreuses et performantes qui les oxydent à grande vitesse par la voie alternative de l'AOX, insensible au cyanure. La voie de la cytochrome C oxydase n'est pas utilisée bien que la cytochrome c oxydase soit détectable. La voie alternative convertit toute (ou presque toute) l'énergie de l'oxydation des coenzymes réduits en chaleur. Fonctionnant à grande vitesse, elle consomme beaucoup d'O2 (crise respiratoire) et libère suffisamment de chaleur pour permettre un échauffement significatif ( la température peut augmenter de 20°C) de l'appendice (phase thermogénique).
Compléments : flexibilité de la chaîne de transfert d'électrons dans la mitochondrie des plantes .