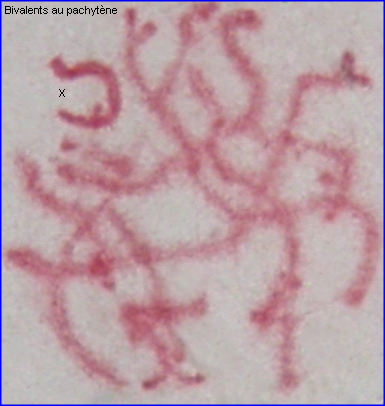
Méiose dans le testicule de Locusta migratoria
Phénomènes chromosomiques à la Prophase I
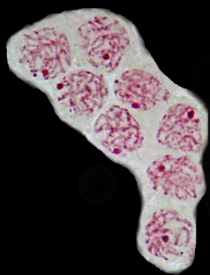
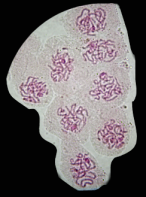
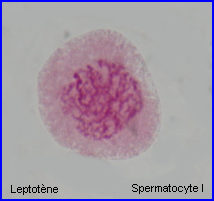
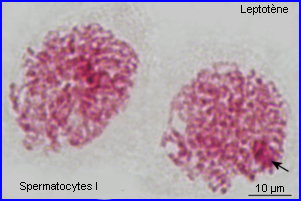
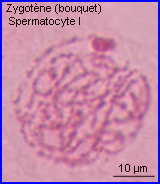
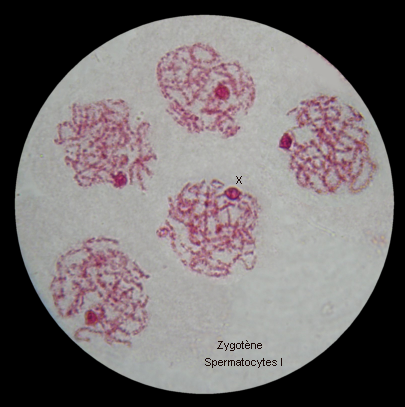
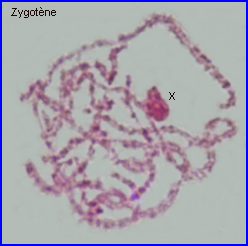

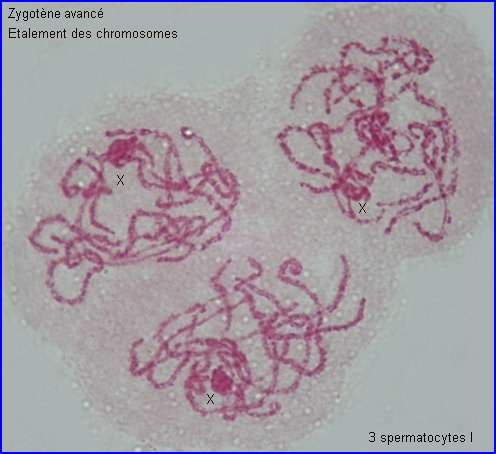
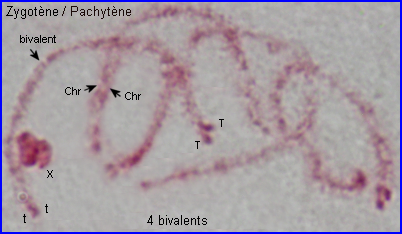
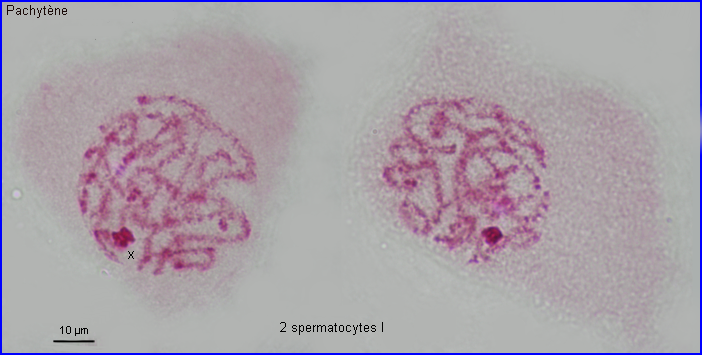
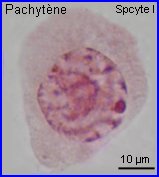
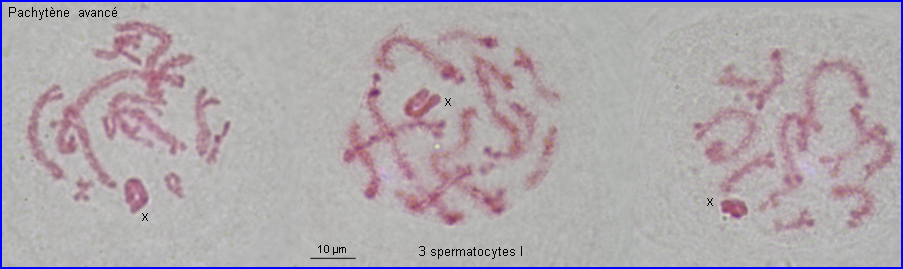
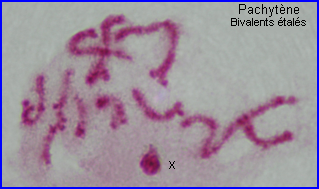
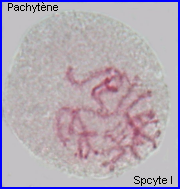
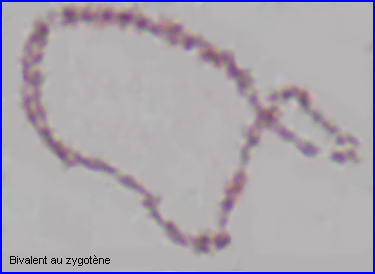
Les extrémités des chromosomes sont redistribuées autour de l'enveloppe nucléaire, la figure en "bouquet" disparaît. Il est parfois possible de compter les bivalents.
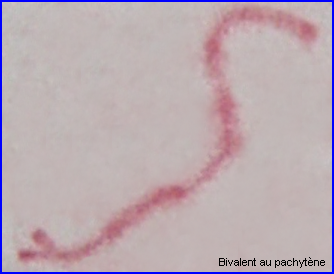
A la transition avec le diplotène, le complexe synaptonémal se désorganise, les homologues se séparent au niveau de chaque bivalent.
Les recombinaisons (résultant des crossing over) qui peuvent déjà se produire au zygotène, ont essentiellement lieu au pachytène.
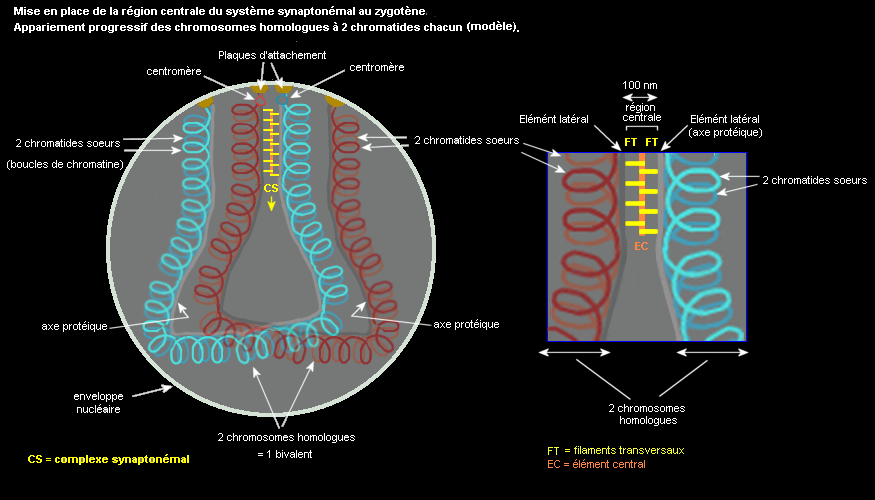
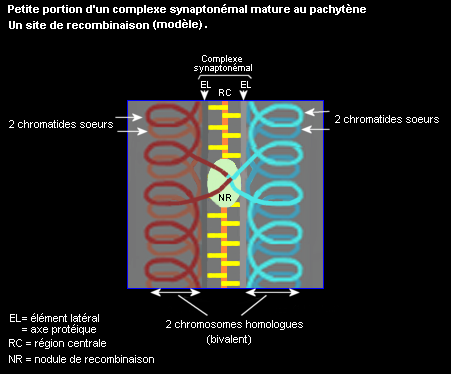
Le complexe synaptonémal, édifice protéique impliqué dans l'appariement des chrosomes homologues et dans les recombinaisons
Un édifice protéique complexe, repérable sur certaines micrographies électroniques réunit les chromosomes homologues à 2 chromatides, c'est le système synaptonémal. Au zygotène, chaque chromosome possède un axe protéique relié à l'enveloppe nucléaire par ses extrémités, dans lequel les boucles de chromatine de chacune des chromatides soeurs sont ancrées. Les axes de chaque homologue se rapprochent et sont solidarisés au cours du zygotène par la mise en place d'un dispositif protéique transversal (central). Au pachytène, le système synaptonémal qui comprend donc les axes (composants latéraux) et l'élément central apparie les chromosomes homologues d'une extrémité à l'autre (bivalents). Certaines protéines du système ont été identifiées, par l'intermédiaire de mutations mais le processus qui permet l'appariement par reconnaissance des parties homologues de chacun des chromosomes d'une paire n'est pas élucidé même si des modèles mettant en jeu les protéines des éléments latéraux ont été proposés. Le pachytène se termine quand les homologues commencent à se séparer par désorganisation du système synaptonémal.
Les recombinaisons génétiques ont lieu au niveau de nodules présents au zygotène et surtout au pachytène et associés à la région centrale du système synaptonémal. Ce sont des complexes multienzymatiques dont l'intervention dans la réalisation des crossing-over a été mise en évidence indirectement. Des protéines qui interviennent dans le cadre des nodules de recombinaison ont été identifiées, le mécanisme moléculaire d'échanges de parties homologues entre chromatides non-soeurs de 2 homologues met en jeu des enzymes qui coupent et d'autres qui soudent l'ADN.