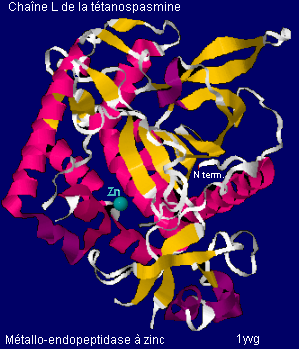
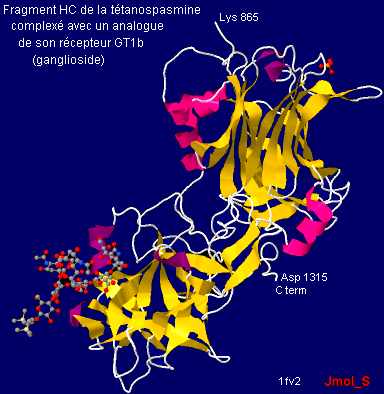
Mécanisme d'action
de la toxine tétanique
La toxine libérée en périphérie, dans la plaie (en milieu anaérobie) par Chlostridium tétani passe dans le milieu intérieur et peut atteindre les terminaisons axoniques des motoneurones alpha. Elle ne franchit pas la barrière hématoencéphalique.
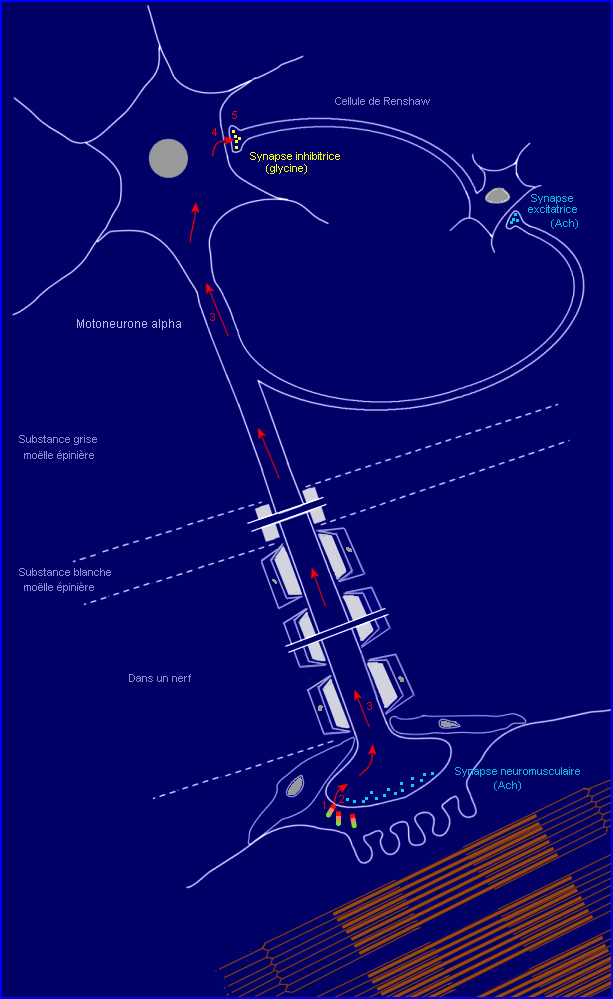
5 : Blocage de l'exocytose du neuromédiateur (glycine) et du fonctionnement de la synapse inhibitrice entre cellule de Renshaw et motoneurone.
4 : Transcytose : libération par le motoneurone (exocytose) puis capture par le neurone de Renshaw (endocytose).
1 : Liaison de la toxine à la membrane d'une terminaison (partie sans myéline) de motoneurone alpha, au niveau d'une synapse neuromusculaire..
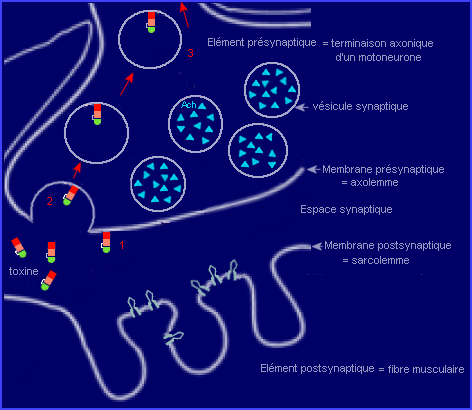
Une molécule de toxine liée à la membrane est ensuite internalisée par endocytose (étape 2) et se retrouve dans une vésicule qui migre dans l'axone du motoneurone, par transport rétrograde (étape 3), vers le corps cellulaire.
- Transcytose de la toxine
La toxine acheminée par transport rétrograde jusque dans le corps cellulaire du motoneurone est libérée par exocytose dans le milieu interstitiel du centre nerveux, la moëlle épinière (étape 4). Elle peut alors se fixer à la membrane plasmique d'une terminaison axonique d'un neurone de Renshaw par l'intermédiaire de son fragment HC qui reconnaît probablement un ganglioside mais aussi une protéine membranaire (étape 5). Les récepteurs qui interviennent ici ne sont peut-être pas les mêmes que ceux mis en jeu lors de la capture périphérique de la toxine par le motoneurone. La toxine fixée est ensuite internalisée par endocytose (étape 6) et se retrouve dans le compartiment endosomique de la cellule de Renshaw.
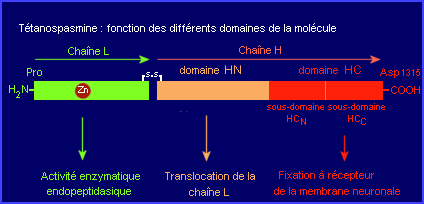
- Translocation de la partie active (toxique) de la toxine du compartiment intravésiculaire vers le cytosol du neurone de Renshaw
La configuration spatiale de la toxine est pH dépendante. L'entrée de protons dans le milieu intravésiculaire (acidification, étape 7) pourrait induire un changement de conformation de la molécule qui deviendrait alors capable par l'intermédiare de son domaine HN de participer (canal transmembranaire) au transfert de sa chaîne L vers le cytosol du neurone. Le processus mis en jeu dans la translocation de la chaîne L (étape 8) reste hypothétique.
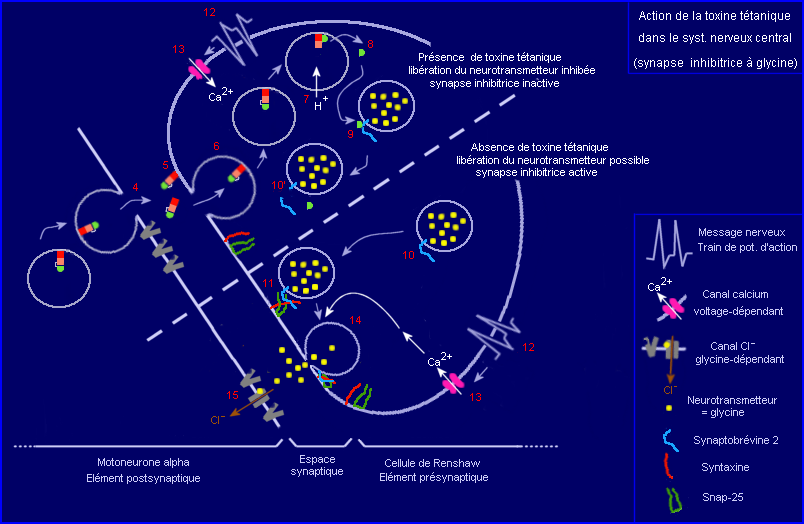
- Action de la partie active de la toxine (chaîne L) sur une molécule cible, la Synaptobrévine 2, impliquée dans l'exocytose du neurotransmetteur glycine.
Pour comprendre le mode d'action de la chaîne L de la toxine au niveau de la synapse inhibitrice glycinergique, proposé par le modèle présenté ci-dessus, il est nécessaire de connaître le rôle central joué par le complexe protéique Synaptobrévine-Syntaxine-Snap 25 dans la transmission synaptique. La fonction de ce complexe apparaît dans la moitié inférieure du schéma, un complément explicatif étant fourni par ailleurs (à voir éventuellement).
Une fois dans le cytosol du neurone de Renshaw, la chaîne L catalyse l'hydrolyse (activité endopeptidasique) d'une liaison peptidique précise de la synaptobrévine 2, petite protéine intégrée à la membrane de la vésicule synaptique (étapes 9 et 10'). Ainsi coupée, la synaptobrévine 2 ne peut plus s'associer avec la syntaxine et snap-25 pour former le complexe (étape 11) impliqué dans le rapprochement vésicule / membrane présynaptique. La formation de ce complexe est indispensable pour permettre ensuite l'exocytose du neurotransmetteur (étape 14), déclenché par le signal Ca2+ (étape 13) dépendant de l'arrivée d'un message nerveux présynaptique (étape 12).
Conséquences physiologiques de l'action de la toxine tétanique
En présence de toxine tétanique (chaîne L), par suite de l'absence de rétrocontrôle inhibiteur, les réponses motrices impliquant les motoneurones alpha peuvent être exacerbées, durables, inadaptées.